

In human populations, changes in genetic variation are driven not only by genetic processes, but can also arise from cultural or social changes. An abrupt population bottleneck specific to human males has been inferred across several Old World (Africa, Europe, Asia) populations 5000–7000 BP. Here, bringing together anthropological theory, recent population genomic studies and mathematical models, we propose a sociocultural hypothesis, involving the formation of patrilineal kin groups and intergroup competition among these groups. Our analysis shows that this sociocultural hypothesis can explain the inference of a population bottleneck. We also show that our hypothesis is consistent with current findings from the archaeogenetics of Old World Eurasia, and is important for conceptions of cultural and social evolution in prehistory. ... If the primary unit of sociopolitical competition is the patrilineal corporate kin group, deaths from intergroup competition, whether in feuds or open warfare, are not randomly distributed, but tend to cluster on the genealogical tree of males. In other words, cultural factors cause biases in the usually random process of transmission of Y-chromosomes, increasing the rate of loss of Y-chromosomal lineages and accelerating genetic drift. Extinction of whole patrilineal groups with common descent would translate to the loss of clades of Y-chromosomes. Furthermore, as success in intergroup competition is associated with group size, borne out empirically in wars [43] as ‘increasing returns at all scales’ [44], and as larger group size may even be associated with increased conflict initiation, borne out in data on feuds45, there may have been positive returns to lineage size. This would accelerate the loss of minor lineages and promote the spread of major ones, further increasing the speed of genetic drift. In addition, the assimilation of women from groups that are disrupted or extirpated through intergroup competition into remaining groups is a common result of warfare in small-scale societies [46]. This, together with female exogamy, would tend to limit the impact of intergroup competition to Y-chromosomes. ... Figure 6 shows a striking pattern of differences in shallowness of coalescence in samples from hunter-gatherer, farmer and pastoralist cultures. While hunter-gatherer Y-chromosomes from the same culture, and often the same sites, commonly divide into haplotypes that coalesce in multiple millennia, Y-chromosomes of samples from farmer and pastoralist cultures are more homogeneous and have more recent coalescences. The Bell Beaker culture has a high proportion of sampled males (81%) from a large geographical area (Iberia to Hungary) who belong to an identical Y-chromosomal haplogroup (R1b-S116), implying common descent from a kin group that existed quite recently. Some groups of males share even more recent descent, on the order of ten generations or fewer [64]. Such recent common descent may even be retained in cultural memory via oral genealogies, such as among descent groups in Northern and Western Africa, whose members can trace descent relationships up to three to four centuries before the generation currently living [40]. Likewise, from Germany to Estonia, the Y-chromosomes of all Corded Ware individuals sampled, except one, belong to a single clade within haplogroup R1a (R1a-M417) and appear to coalesce shortly before sample deposition.Citation... Zeng et al., Cultural hitchhiking and competition between patrilineal kin groups explain the post-Neolithic Y-chromosome bottleneck, Nature Communicationsvolume 9, Article number: 2077 (2018) doi:10.1038/s41467-018-04375-6 Update 30/05/2018: For those clued in, here's an awesome quote from the relevant press release.Thus, groups of males in European post-Neolithic agropastoralist cultures appear to descend patrilineally from a comparatively smaller number of progenitors when compared to hunter gatherers, and this pattern is especially pronounced among pastoralists. Our hypothesis would predict that post-Neolithic societies, despite their larger population size, have difficulty retaining ancestral diversity of Y-chromosomes due to mechanisms that accelerate their genetic drift, which is certainly in accord with the data. The tendency of pastoralist cultures to show the lowest Y-chromosomal diversity and the shallowest coalescence would also be explained, as they may have experienced the social conditions that characterized cultures of the Central Asian steppes [42]. Indeed, the Corded Ware pastoralists may have been organized into segmentary lineages [65], an extremely common tribal system among pastoralist cultures, including those of historical Central Asia [66].
The outlines of that idea came to Tian Chen Zeng, a Stanford undergraduate in sociology, after spending hours reading blog posts that speculated - unconvincingly, Zeng thought - on the origins of the "Neolithic Y-chromosome bottleneck," as the event is known.See also... Late PIE ground zero now obvious; location of PIE homeland still uncertain, but...

The indigenous populations of inner Eurasia, a huge geographic region covering the central Eurasian steppe and the northern Eurasian taiga and tundra, harbor tremendous diversity in their genes, cultures and languages. In this study, we report novel genome-wide data for 763 individuals from Armenia, Georgia, Kazakhstan, Moldova, Mongolia, Russia, Tajikistan, Ukraine, and Uzbekistan. We furthermore report genome-wide data of two Eneolithic individuals (~5,400 years before present) associated with the Botai culture in northern Kazakhstan. We find that inner Eurasian populations are structured into three distinct admixture clines stretching between various western and eastern Eurasian ancestries. This genetic separation is well mirrored by geography. The ancient Botai genomes suggest yet another layer of admixture in inner Eurasia that involves Mesolithic hunter-gatherers in Europe, the Upper Paleolithic southern Siberians and East Asians. Admixture modeling of ancient and modern populations suggests an overwriting of this ancient structure in the Altai-Sayan region by migrations of western steppe herders, but partial retaining of this ancient North Eurasian-related cline further to the North. Finally, the genetic structure of Caucasus populations highlights a role of the Caucasus Mountains as a barrier to gene flow and suggests a post-Neolithic gene flow into North Caucasus populations from the steppe.

G25_Europe_scaled.datYou should end up with the plot below. Note that to see the group labels and outlines, you need to tick the appropriate boxes in the panel to the right of the image. To improve the experience, it might also be useful to color-code different parts of Europe, and you can do that by choosing Edit > Row colors/symbols. Of course, if you have Global25 coordinates you can add yourself to the datasheet to see where you plot.


G25_West_Eurasia_scaled.datDownload the datasheet, plug it into PAST, select all of the columns by clicking on the empty cell above the labels, and go to Multivariate > Ordination > Principal Components. Here's a screen cap of me doing it:



Based on PCA and ADMIXTURE plots we observe two distinct genetic clusters: one cluster falls with previously published ancient individuals from the West Eurasian steppe (hence termed ‘Steppe’), and the second clusters with present-day southern Caucasian populations and ancient Bronze Age individuals from today’s Armenia (henceforth called ‘Caucasus’), while a few individuals take on intermediate positions between the two. The stark distinction seen in our temporal transect is also visible in the Y-chromosome haplogroup distribution, with R1/R1b1 and Q1a2 types in the Steppe and L, J, and G2 types in the Caucasus cluster (Fig. 3A, Supplementary Data 1). In contrast, the mitochondrial haplogroup distribution is more diverse and almost identical in both groups (Fig. 3B, Supplementary Data 1).

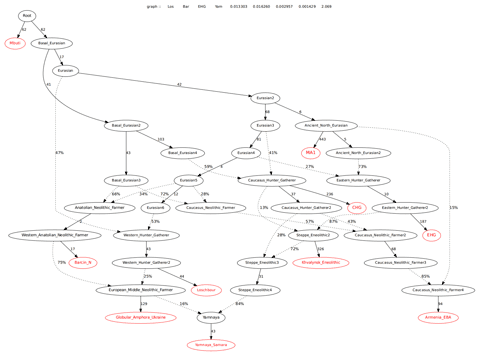
Importantly, our results show a subtle contribution of both Anatolian farmer-related ancestry and WHG-related ancestry (Fig.4; Supplementary Tables 13 and 14), which was likely contributed through Middle and Late Neolithic farming groups from adjacent regions in the West. A direct source of Anatolian farmer-related ancestry can be ruled out (Supplementary Table 15). At present, due to the limits of our resolution, we cannot identify a single best source population. However, geographically proximal and contemporaneous groups such as Globular Amphora and Eneolithic groups from the Black Sea area (Ukraine and Bulgaria), which represent all four distal sources (CHG, EHG, WHG, and Anatolian_Neolithic) are among the best supported candidates (Fig. 4; Supplementary Tables 13,14 and 15).Check out what I had to say about this issue exactly two years ago: Yamnaya = Khvalynsk + extra CHG + maybe something else. Not bragging, just making a point that I do know what I'm doing here, most of the time anyway. Wang et al. conclude their preprint with, unfortunately I have to say, some downright bizarre comments in regards to the Proto-Indo-European (PIE) homeland debate. But I'll get back to that later, when the ancient data from this and forthcoming related papers are released online. Citation... Wang et al., The genetic prehistory of the Greater Caucasus, bioRxiv, posted May 16, 2018, doi: https://doi.org/10.1101/322347 See also... Yamnaya: home-grown Ahead of the pack Genetic borders are usually linguistic borders too

Global25 datasheet ancient scaled Global25 pop averages ancient scaled Global25 datasheet ancient Global25 pop averages ancientThe interesting thing about those Tian Shan nomads, especially the Kangju people, is that they're much more West Eurasian (European + West Asian) than the Asian Scythians sampled to date. However, despite this, they're still no good for modeling the West Eurasian ancestry of most South Asian populations. I've looked at this closely, and the Steppe_MLBA cluster is still the one to beat in this respect. See also... Genetic ancestry online store (to be updated regularly)
Anatolia_MLBA_MA2203 Anatolia_EBA 0.794±0.073 Ukraine_Eneolithic_I6561 0.206±0.073 tail: 0.400704 Full outputObviously, these numbers aren't exactly impressive. But if the signal is real, then it might be an indication of things to come when someone manages to sequence at least a few genomes from confirmed Hittite remains. None of the other Anatolia_MLBA individuals, three of whom are from the Assyrian Colony Period (~2000–1750 BCE), show such obvious steppe ancestry.
Anatolia_MLBA_MA2200 Anatolia_EBA 1.000 Ukraine_Eneolithic_I6561 0.000 tail: 0.449485 Full output Anatolia_MLBA_MA2205 Anatolia_EBA 0.983±0.069 Ukraine_Eneolithic_I6561 0.017±0.069 tail: 0.618499 Full output Anatolia_MLBA_MA2206 Anatolia_EBA 0.868±0.089 Ukraine_Eneolithic_I6561 0.132±0.089 tail: 0.708811 Full output Anatolia_MLBA w/o MA2203 Anatolia_EBA 1.000 Ukraine_Eneolithic_I6561 0.000 tail: 0.286377 Full outputIn any case, apart from all of that, Damgaard et al. do take a measured and sober approach to interpreting their archaeogenetic data in the context of the Indo-European homeland debate. The paper also includes a very thorough linguistic supplement, freely available here, which reveals that there is Eastern European Hunter-Gatherer (EHG) ancestry in soon to be published Maykop culture samples. From the supplement (emphasis is mine):
Despite a general agreement on a Pontic-Caspian origin of the Anatolian Indo-European language family, it is currently impossible to determine on linguistic grounds whether the language reached Anatolia through the Balkans in the West (Anthony 2007; Mallory 1989: 30; Melchert 2003; Steiner 1990; Watkins 2006: 50) or through the Caucasus in the East (Kristiansen 2005: 77; Stefanini 2002; Winn 1981). From their earliest attestations, the Anatolian languages are clustered in Anatolia, and if the distribution reflects a prehistoric linguistic speciation event (as argued by Oettinger 2002: 52), then it may be taken as an indication that the arrival and disintegration of Proto-Anatolian language took place in the same area (Steiner 1981: 169). However, others have reasoned that the estimated period between the dissolution of the Proto-Anatolian language and the attestation of the individual daughter languages is extensive enough to allow for prehistoric mobility within Anatolia, theoretically leaving plenty of time for secondary East-to-West dispersals (cf. Melchert 2003: 25). Whatever the case may be, there are no linguistic indications for any mass migration of steppe-derived Anatolian speakers dominating or replacing local populations. Rather, the Anatolian Indo-European languages appear in history as an organically integrated part of the linguistic landscape. In lexicon, syntax, and phonology, the second millennium languages of Anatolia formed a convergent, diffusional linguistic area (Watkins 2001: 54). Though the presence of an Indo-European language itself demonstrates that a certain number of speakers must have entered the area, the establishment of the Anatolian Indo-European branch in Anatolia is likely to have happened through a long-term process of infiltration and acculturalization rather than through mass immigration or elite dominance (Melchert 2003: 25). Furthermore, the genetic results presented in Damgaard et al. 2018 show no indication of a large-scale intrusion of a steppe population. The EHG ancestry detected in individuals associated with both Yamnaya (3000–2400 BCE) and the Maykop culture (3700–3000 BCE) (in prep.) is absent from our Anatolian specimens, suggesting that neither archaeological horizon constitutes a suitable candidate for a “homeland” or “stepping stone” for the origin or spread of Anatolian Indo-European speakers to Anatolia. However, with the archaeological and genetic data presented here, we cannot reject a continuous small-scale influx of mixed groups from the direction of the Caucasus during the Chalcolithic period of the 4th millennium BCE. ... Under the “Steppe Hypothesis,” the Indo-Iranian languages are not seen as indigenous to South Asia but rather as an intrusive branch from the northern steppe zone (cf. Anthony 2007: 408–411; Mallory 1989: 35–56; Parpola 1995; Witzel 1999, 2001). Important clues to the original location and dispersal of the Indo-Iranians into South and Southwest Asia are provided by the Indo-Iranian languages themselves. The Indo-Aryan and Iranian languages share a common set of etymologically related terms related to equestrianism and chariotry (Malandra 1991). Since it can be shown that this terminology was inherited from their Proto-Indo-Iranian ancestor, rather than independently borrowed from a third language, the split of this ancestor into Indo-Aryan and Iranian languages must postdate these technological innovations. The earliest available archaeological evidence of two-wheeled chariots is dated to approximately 2000 BCE (Anthony 1995; Anthony and Ringe 2015; Kuznetsov 2006: 638–645; Teufer 2012: 282). This offers the earliest possible date so far for the end of Proto-Indo-Iranian as a linguistic unity. The reference to a mariannu in a text from Tell speakers. Leilān in Syria discussed below pushes the latest possible period of Indo-Iranian linguistic unity to the 18th century BCE. ... The traces of early Indo-Aryan speakers in Northern Syria positions the oldest Indo-Iranian speakers somewhere between Western Asia and the Greater Punjab, where the earliest Vedic text is thought to have been composed during the Late Bronze Age (cf. Witzel 1999: 3). In addition, a northern connection is suggested by contacts between the Indo-Iranian and the Finno-Ugric languages. Speakers of the Finno-Ugric family, whose antecedent is commonly sought in the vicinity of the Ural Mountains, followed an east-to-west trajectory through the forest zone north and directly adjacent to the steppes, producing languages across to the Baltic Sea. In the languages that split off along this trajectory, loanwords from various stages in the development of the Indo-Iranian languages can be distinguished: 1) Pre-Proto-Indo-Iranian (Proto-Finno-Ugric *kekrä (cycle), *kesträ (spindle), and *-teksä (ten) are borrowed from early preforms of Sanskrit cakrá- (wheel, cycle), cattra- (spindle), and daśa- (10); Koivulehto 2001), 2) Proto-Indo-Iranian (Proto-Finno-Ugric *śata (one hundred) is borrowed from a form close to Sanskrit śatám (one hundred), 3) Pre-Proto-Indo-Aryan (Proto-Finno-Ugric *ora (awl), *reśmä (rope), and *ant- (young grass) are borrowed from preforms of Sanskrit ā́ r ā- (awl), raśmí- (rein), and ándhas- (grass); Koivulehto 2001: 250; Lubotsky 2001: 308), and 4) loanwords from later stages of Iranian (Koivulehto 2001; Korenchy 1972). The period of prehistoric language contact with Finno-Ugric thus covers the entire evolution of Pre-Proto-Indo-Iranian into Proto-Indo-Iranian, as well as the dissolution of the latter into Proto-Indo-Aryan and Proto-Iranian. As such, it situates the prehistoric location of the Indo-Iranian branch around the southern Urals (Kuz’mina 2001).Citation... Guus Kroonen, Gojko Barjamovic, & Michaël Peyrot. (2018). Linguistic supplement to Damgaard et al. 2018: Early Indo-European languages, Anatolian, Tocharian and Indo-Iranian. http://doi.org/10.5281/zenodo.1240524 Update 14/05/2018: I managed to, more or less, reproduce my qpAdm models with qpGraph. This is never a simple and easy task, so I'm now more confident that Anatolia_MLBA MA2203 really does harbor ancestry from the steppe.












SPGT Indus_Periphery 0.692±0.042 Shahr_I_Sokhta_BA1 0.104±0.045 Sintashta_MLBA 0.204±0.015 taildiff: 0.659609 Full outputI'm trying to incorporate this new information into my Admixture graph models of the SPGT groups (see here). If I manage to come up with something useful I'll update this post with the results. Update 08/05/2018: see here.
Ancient Central and South Asia genetic distance (Fst) matrixHint: it's probably easiest to initially explore this format with a program called PAST. Indeed, if you'd like to model fine scale ancestry proportions based on these data, it might be a good idea to use PAST to first turn the matrix into a principal coordinates (PCoA) datasheet (like this). On a related note, as I was typing this, commentator Chetan alerted me to a post at the Molgen forum claiming that Y-haplogroup R1b-L51 has turned up in Eneolithic remains from Pontic-Caspian steppe (see here). If true, then it's a big deal, because it's the best evidence yet that L51 expanded into Central and Western Europe from the steppes. This is the Google translation of the post. Emphasis is mine.
Hello. Today, the XIV Samara Archeological Conference was held. The following reports were heard. Khokhlov AA Preliminary results of anthropological and genetic studies of materials of the Volga-Ural region of the Neolithic-Early Bronze Age by an international group of scientists. In his report, AA Khokhlov. introduced into scientific circulation until the unpublished data of the new Eneolithic burial ground Ekatirinovsky cape, which combines both the Mariupol and Khvalyn features, and refers to the fourth quarter of the V millennium BC. All samples analyzed had a uraloid anthropological type, the chromosome of all the samples belonged to the haplogroup R1b1a2 (R-P 312 / S 116), and the haplogroup R1b1a1a2a1a1c2b2b1a2. Mito to haplogroups U2, U4, U5. In the Khvalyn burial grounds (1 half of the 4th millennium BC), the anthropological material differs in a greater variety. In addition to the uraloid substratum, European broad-leaved and southern-European variants are recorded. To the game haplogroup R1a1, O1a1, I2a2 are added to mito T2a1b, H2a1.I'd say that this information sounds legit. But let's wait and see if the results are backed up by one of the major ancient DNA labs in the west, like the Reich Lab. See also... Late PIE ground zero now obvious; location of PIE homeland still uncertain, but...

{kind=link}