The earliest ancient DNA data of modern humans from Europe dates to ~40 thousand years ago, but that from the Caucasus and the Near East to only ~14 thousand years ago, from populations who lived long after the Last Glacial Maximum (LGM) ~26.5-19 thousand years ago. To address this imbalance and to better understand the relationship of Europeans and Near Easterners, we report genome-wide data from two ~26 thousand year old individuals from Dzudzuana Cave in Georgia in the Caucasus from around the beginning of the LGM. Surprisingly, the Dzudzuana population was more closely related to early agriculturalists from western Anatolia ~8 thousand years ago than to the hunter-gatherers of the Caucasus from the same region of western Georgia of ~13-10 thousand years ago. Most of the Dzudzuana population's ancestry was deeply related to the post-glacial western European hunter-gatherers of the 'Villabruna cluster', but it also had ancestry from a lineage that had separated from the great majority of non-African populations before they separated from each other, proving that such 'Basal Eurasians' were present in West Eurasia twice as early as previously recorded. We document major population turnover in the Near East after the time of Dzudzuana, showing that the highly differentiated Holocene populations of the region were formed by 'Ancient North Eurasian' admixture into the Caucasus and Iran and North African admixture into the Natufians of the Levant. We finally show that the Dzudzuana population contributed the majority of the ancestry of post-Ice Age people in the Near East, North Africa, and even parts of Europe, thereby becoming the largest single contributor of ancestry of all present-day West Eurasians.Lazaridis et al., Paleolithic DNA from the Caucasus reveals core of West Eurasian ancestry, bioRxiv, posted September 21, 2018, doi: https://doi.org/10.1101/423079 See also... Villabruna cluster =/= Near Eastern migrants
Friday, September 21, 2018
Dzudzuana Ice Age foragers: a different type of Caucasus hunter-gatherer (Lazaridis et al. 2018 preprint)
Over at bioRxiv at this LINK. Below is the abstract. Emphasis is mine.
Thursday, September 20, 2018
Early Anatolian farmers were overwhelmingly of local hunter-gatherer origin (Feldman et al. 2018 preprint)
Over at bioRxiv at this LINK. The dataset in this preprint includes just one Anatolian hunter-gatherer, but that's enough to make the point that in Anatolia, unlike in Europe, there was very strong genetic continuity between the local foragers and earliest farmers. His Y-chromosome haplogroup is an interesting one: C1a2, which has been recorded in European remains from the Upper Paleolithic. Below is the abstract and a pertinent quote. I think this preprint basically confirms what I argued about the origin of the so called Villabruna hunter-gatherer clade back in 2016 (see here). Emphasis is mine.
Anatolia was home to some of the earliest farming communities. It has been long debated whether a migration of farming groups introduced agriculture to central Anatolia. Here, we report the first genome-wide data from a 15,000 year-old Anatolian hunter-gatherer and from seven Anatolian and Levantine early farmers. We find high genetic continuity between the hunter-gatherer and early farmers of Anatolia and detect two distinct incoming ancestries: an early Iranian/Caucasus related one and a later one linked to the ancient Levant. Finally, we observe a genetic link between southern Europe and the Near East predating 15,000 years ago that extends to central Europe during the post-last-glacial maximum period. Our results suggest a limited role of human migration in the emergence of agriculture in central Anatolia. ... Among the Later European HG, recently reported Mesolithic hunter-gatherers from the Balkan peninsula, which geographically connects Anatolia and central Europe (‘Iron Gates HG’) [18], are genetically closer to AHG when compared to all the other European hunter-gatherers, as shown in the significantly positive statistic D(Iron_Gates_HG, European hunter-gatherers; AHG, Mbuti/Altai). Iron Gates HG are followed by Epigravettian and Mesolithic individuals from Italy and France (Villabruna [14] and Ranchot88 respectively [17]) as the next two European hunter-gatherers genetically closest to AHG [20] (Fig. 3A and data table S5). Iron Gates HG have been suggested to be genetically intermediate between WHG and eastern European hunter-gatherers (EHG) with an additional unknown ancestral component [18]. We find that Iron Gates HG can be modeled as a three-way mixture of Near-Eastern hunter-gatherers (25.8 ± 5.0 % AHG or 11.1 ± 2.2 % Natufian), WHG (62.9 ± 7.4 % or 78.0 ± 4.6 % respectively) and EHG (11.3 ± 3.3 % or 10.9 ± 3 % respectively); (tables S4 and S9). The affinity detected by the above D-statistic can be explained by gene flow from Near-Eastern hunter-gatherers into the ancestors of Iron Gates or by a gene flow from a population ancestral to Iron Gates into the Near-Eastern hunter-gatherers as well as by a combination of both. To distinguish the direction of the gene flow, we examined the Basal Eurasian ancestry component (α), which is prevalent in the Near East [6] but undetectable in European hunter-gatherers [17]. Following a published approach [6], we estimated α to be 24.8 ± 5.5 % in AHG and 38.5 ± 5.0 % in Natufians (Fig. 3B, table S10), consistent with previous estimates for the latter [6]. Under the model of unidirectional gene flow from Anatolia to Europe, 6.4 % is expected for α of Iron Gates by calculating (% AHG in Iron Gates HG) × (α in AHG). However, Iron Gates can be modeled without any Basal Eurasian ancestry or with a non-significant proportion of 1.6 ± 2.8 % (Fig. 3B, table S10), suggesting that unidirectional gene flow from the Near East to Europe alone is insufficient to explain the extra affinity between the Iron Gates HG and the Near-Eastern hunter-gatherers. Thus, it is plausible to assume that prior to 15,000 years ago there was either a bidirectional gene flow between populations ancestral to Southeastern Europeans of the early Holocene and Anatolians of the late glacial or a dispersal of Southeastern Europeans into the Near East. Presumably, this Southeastern European ancestral population later spread into central Europe during the post-last-glacial maximum (LGM) period, resulting in the observed late Pleistocene genetic affinity between the Near East and Europe.Feldman et al., Late Pleistocene human genome suggests a local origin for the first farmers of central Anatolia, biRxiv, posted September 20, 2018, doi: https://doi.org/10.1101/422295
Sunday, September 16, 2018
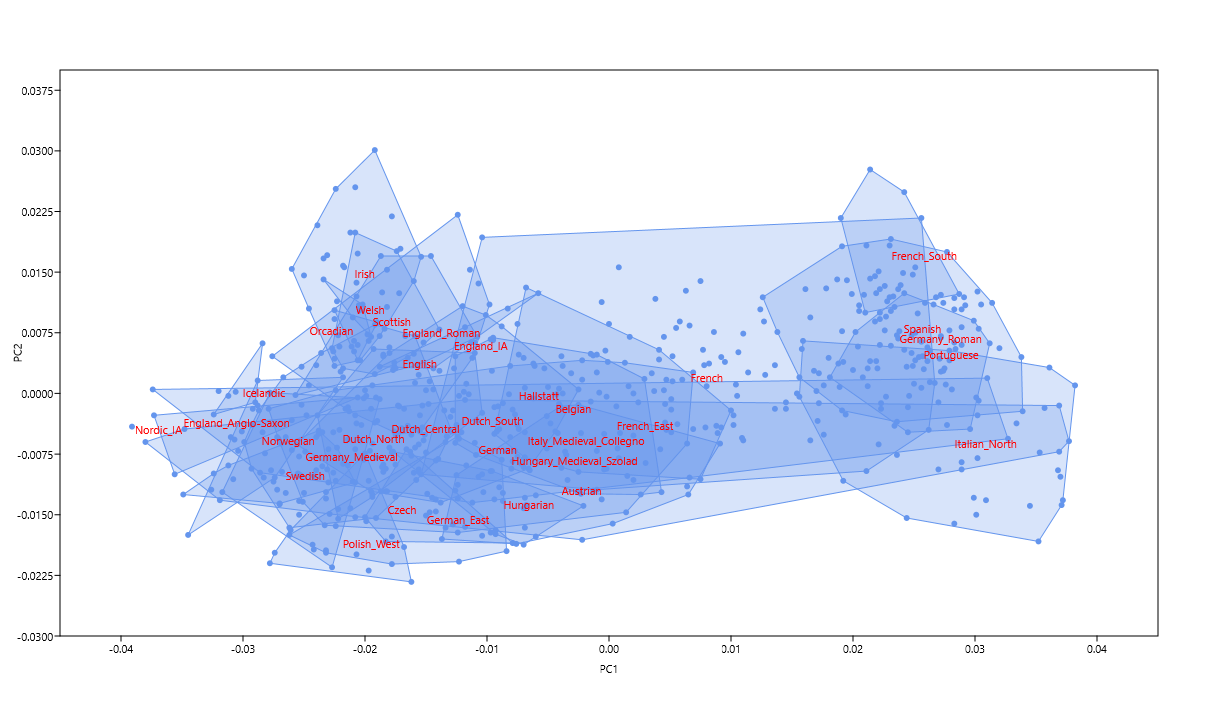
Celtic vs Germanic Europe
I have a feeling that ancient DNA from post-Bronze Age Northwestern Europe will be coming thick and fast from now on. To get the most out of such data I've designed a new Principal Component Analysis (PCA) that does a better job of separating the Celtic- and Germanic-speaking populations of Europe than my previous efforts of this sort (see here and here). Below are two different versions of the same PCA. The relevant datasheet is available here.

 And here's a Discrimination Analysis (LDA) plot based on the 25 principal components. It further differentiates many of the populations along the east > west cline of genetic diversity.
And here's a Discrimination Analysis (LDA) plot based on the 25 principal components. It further differentiates many of the populations along the east > west cline of genetic diversity.
 The difference between the Germanic Anglo-Saxons and the Celtic and Roman Britons of what is now eastern England is obvious. The Anglo-Saxons could pass for Scandinavians, while the Celts and Romans both cluster between the Irish and French. This makes good sense, and is exactly what I was looking for. It's also interesting to see the presumably Celtic-speaking Hallstatt samples from Bylany, Czechia, clustering with the Belgians.
Update 14/12/2019: Pictured below is a new version of my Celtic vs Germanic genetic map. It's based on the same Principal Component Analysis (PCA) as the original, but more focused on Northwestern Europe and produced with a different program.
The difference between the Germanic Anglo-Saxons and the Celtic and Roman Britons of what is now eastern England is obvious. The Anglo-Saxons could pass for Scandinavians, while the Celts and Romans both cluster between the Irish and French. This makes good sense, and is exactly what I was looking for. It's also interesting to see the presumably Celtic-speaking Hallstatt samples from Bylany, Czechia, clustering with the Belgians.
Update 14/12/2019: Pictured below is a new version of my Celtic vs Germanic genetic map. It's based on the same Principal Component Analysis (PCA) as the original, but more focused on Northwestern Europe and produced with a different program.
 To see the interactive online version, navigate to Vahaduo Custom PCA and copy paste the text from here into the empty space under the PCA DATA tab. Then press the PLOT PCA button under the PCA PLOT tab. For more guidance, refer to the screen caps here and here.
To include a wider range of populations in the key, just edit the data accordingly. For instance, to break up the ancient grouping into more specific populations, delete the Ancient: prefix in all of the relevant rows. This is what you should see:
To see the interactive online version, navigate to Vahaduo Custom PCA and copy paste the text from here into the empty space under the PCA DATA tab. Then press the PLOT PCA button under the PCA PLOT tab. For more guidance, refer to the screen caps here and here.
To include a wider range of populations in the key, just edit the data accordingly. For instance, to break up the ancient grouping into more specific populations, delete the Ancient: prefix in all of the relevant rows. This is what you should see:
 Conversely, you can leave the ancient sample set intact and instead reorder the present-day linguistic groupings into, say, geographic groupings. To achieve this just delete all of the linguistic prefixes, such as Celtic:, Germanic:, and so on. You should end up with a datasheet like this and plot like this.
Of course, you can design your own plot by using any combination of the ancient and present-day individuals and populations that I've already run in this PCA. Their coordinates are listed here. Indeed, if you're in the possession of your own Celtic vs Germanic PCA coordinates, you can add yourself to the plot. And if you're not, see here.
It's also possible to re-process PCA data via the SOURCE tab. But I don't recommend doing this with the Celtic vs Germanic data, which are derived from a fine scale analysis and don't pack much variation. On the other hand, Global25 data are ideal for such re-processing. I made the plots below from subsets of Global25 coordinates available in a zip file here. To see how, refer to the screen caps here and here.
Conversely, you can leave the ancient sample set intact and instead reorder the present-day linguistic groupings into, say, geographic groupings. To achieve this just delete all of the linguistic prefixes, such as Celtic:, Germanic:, and so on. You should end up with a datasheet like this and plot like this.
Of course, you can design your own plot by using any combination of the ancient and present-day individuals and populations that I've already run in this PCA. Their coordinates are listed here. Indeed, if you're in the possession of your own Celtic vs Germanic PCA coordinates, you can add yourself to the plot. And if you're not, see here.
It's also possible to re-process PCA data via the SOURCE tab. But I don't recommend doing this with the Celtic vs Germanic data, which are derived from a fine scale analysis and don't pack much variation. On the other hand, Global25 data are ideal for such re-processing. I made the plots below from subsets of Global25 coordinates available in a zip file here. To see how, refer to the screen caps here and here.


 See also...
Modeling your ancestry has never been easier
Getting the most out of the Global25
Modeling genetic ancestry with Davidski: step by step
See also...
Modeling your ancestry has never been easier
Getting the most out of the Global25
Modeling genetic ancestry with Davidski: step by step



{kind=link}
{kind=link}

{kind=link}
{kind=link}
{kind=link}



Wednesday, September 12, 2018
Avars and Longobards
Most of the "barbarians" from today's Amorim et al. paper have made it into the Global25 datasheets. Look for the samples with Collegno and Szolad in their labels. Same links as always...
 And this is my Global25-derived North European PCA featuring a subset of these samples that plotted firmly with present-day populations from north of the Alps, Balkans and Pyrenees. The aforementioned Avars (red dots) are sitting within the Polish cluster. The relevant datasheet is available here.
And this is my Global25-derived North European PCA featuring a subset of these samples that plotted firmly with present-day populations from north of the Alps, Balkans and Pyrenees. The aforementioned Avars (red dots) are sitting within the Polish cluster. The relevant datasheet is available here.
 See also...
Greeks in a Longobard cemetery
First real foray into Migration Period Europe: the Gepid, Roman, Ostrogoth and others
See also...
Greeks in a Longobard cemetery
First real foray into Migration Period Europe: the Gepid, Roman, Ostrogoth and others
Global25 datasheet ancient scaled Global25 pop averages ancient scaled Global25 datasheet ancient Global25 pop averages ancientHere's my usual Principal Component Analysis (PCA) of West Eurasian variation with the same individuals. As seen in the paper, the two females from Avar burials are very European indeed, with no hints of any recent Asian ancestry. The relevant datasheet is available here.


Tuesday, September 11, 2018
Blast from the past: Yamnaya prediction from 2016
I wonder what's holding up the publication of the Wang et al. "Greater Caucasus" preprint? It was released back in May at the bioRxiv (see here). On a related note, I was looking back at some of the stuff that I wrote about the origin of the Yamnaya people (aka Steppe_EMBA), and found this...
But here's my prediction: Steppe_EMBA only has 10-15% admixture from the post-Mesolithic Near East not including the North Caucasus, and basically all of this comes via female mediated gene flow from farming communities in the Caucasus and perhaps present-day Ukraine.The relevant blog post from 2016 is here. I totally forgot that I made such a bold prediction. But it actually has a very good chance now of being proven correct, more or less. This, however, depends on the precise origin of the Yamnaya-like Eneolithic populations of the southernmost parts of the Pontic-Caspian steppe. But, considering the data in Wang et al., I think the possibility that they date back to the Pottery Neolithic period, and are thus indigenous to the region, looks quite high. About a year later I made a prediction about the genetic structure of the Maykop people, and was basically proven right by Wang et al. (see here). Admittedly, my jaw dropped when I saw how the Steppe Maykop individuals came out in the preprint, with their Botai-like ancestry that is missing in all Yamnaya populations sampled to date. But it was an interesting outcome and nice to be surprised by ancient DNA yet again. See also... Genetic borders are usually linguistic borders too Ahead of the pack Open thread: What are the linguistic implications of Olalde et al. 2019?
Wednesday, September 5, 2018
ISBA 2018 abstracts
The ISBA 2018 conference is in a couple of weeks and the abstract book is now available here. Below are a few examples of what's on offer this year. Admittedly, the Scythian abstract looks a bit weird to me, because we know for a fact that the Scythians who lived in the Pontic-Caspian steppe harbored Siberian genome-wide and maternal admixture (see here and here). The abstract about the horses and mules looks like it's from the major horse paper that I blogged about a few days ago (see here). Emphasis is mine...
Genetic continuity in the western Eurasian Steppe broken not due to Scythian dominance, but rather at the transition to the Chernyakhov culture (Ostrogoths) Jarve et al. The long-held archaeological view sees the Early Iron Age nomadic Scythians expanding west from their Altai region homeland across the Eurasian Steppe until they reached the Ponto-Caspian region north of the Black and Caspian Seas by around 2,900 BP. However, the migration theory has not found support from ancient DNA evidence, and it is still unclear how much of the Scythian dominance in the Eurasian Steppe was due to movements of people and how much reflected cultural diffusion and elite dominance. We present new whole-genome results of 31 ancient Western and Eastern Scythians as well as samples pre- and postdating them that allow us to set the Scythians in a temporal context by comparing the Western Scythians to samples before and after within the Ponto-Caspian region. We detect no significant contribution of the Scythians to the Early Iron Age Ponto-Caspian gene pool, inferring instead a genetic continuity in the western Eurasian Steppe that persisted from at least 4,800–4,400 cal BP to 2,700–2,100 cal BP (based on our radiocarbon dated samples), i.e. from the Yamnaya through the Scythian period. However, the transition from the Scythian to the Chernyakhov culture between 2,100 and 1,700 cal BP does mark a shift in the Ponto-Caspian genetic landscape, with various analyses showing that Chernyakhov culture samples share more drift and derived alleles with Bronze/Iron Age and modern Europeans, while the Scythians position outside modern European variation. Our results agree well with the Ostrogothic origins of the Chernyakhov culture and support the hypothesis that the Scythian dominance was cultural rather than achieved through population replacement. ... Unveiling early horse domestication and mule production with ancient genome-scale data Fages et al. Despite being one of the last large herbivores to be domesticated, the horse has deeply transformed human civilizations. It provided not only important primary domestication products including both meat and milk, but also invaluable secondary products, such as fast transportation, which impacted patterns of human movements and facilitated the spread of vast cultural and political units across the Old World. The steps underpinning early horse domestication are, however, difficult to track in the archaeological record, especially due to (1) the relative scarcity of horse bone assemblages until the Neolithic and Bronze Age transition, and (2) the absence of clear patterns of size differentiation prior to the Iron Age. Some of the more recent steps accompanying horse domestication, and in particular how it was transformed to fit a range of utilizations in different human groups, are also poorly documented. One such step pertains to the development of mules, and other kinds of F1-hybrids, which are difficult to identify on fragmentary remains using morphology alone. Within the course of the ERC PEGASUS project, we have generated genome-scale sequence information from hundreds of equine archaeological remains spread across Eurasia and spanning the last ~40,000 years. These data helped us test the extent to which candidate domestication centres in Central Asia and Europe contributed to the genetic makeup of the modern domestic horse and propose a minimal time boundary for the earliest utilization of mules by mankind. ... The genetic history of the Iberian Peninsula over the last 8000 years Olalde et al. The Iberian Peninsula, lying on the southwestern corner of Europe, provides an excellent opportunity to assess the final impact of population movements entering the continent from the east and to study prehistoric and historic connections with North Africa. Previous studies have addressed the population history of Iberia using ancient genomes, but the final steps leading to the formation of the modern Iberian gene pool during the last 4000 years remain largely unexplored. Here we report genome-wide data from 153 ancient individuals from Iberia, more than doubling the number of available genomes from this region and providing the most comprehensive genetic transect of any region in the world during the last 8000 years. We find that Mesolithic hunter-gatherers dated to the last centuries before the arrival of farmers showed an increased genetic affinity to central European hunter-gatherers, as compared to earlier individuals. During the third millennium BCE, Iberia received newcomers from south and north. The presence of one individual with a North African origin in central Iberia demonstrates early sporadic contacts across the strait of Gibraltar. Beginning ~2500 BCE, the arrival of individuals with steppe-related ancestry had a rapid and widespread genetic impact, with Bronze Age populations deriving ~40% of their autosomal ancestry and 100% of their Y-chromosomes from these migrants. During the later Iron Age, the first genome-wide data from ancient non-Indo-European speakers showed that they were similar to contemporaneous Indo-European speakers and derived most of their ancestry from the earlier Bronze Age substratum. With the exception of Basques, who remain broadly similar to Iron Age populations, during the last 2500 years Iberian populations were affected by additional gene-flow from the Central/Eastern Mediterranean region, probably associated to the Roman conquest, and from North Africa during the Moorish conquest but also in earlier periods, probably related to the Phoenician-Punic colonization of Southern Iberia.See also... How relevant is Arslantepe to the PIE homeland debate?
Sunday, September 2, 2018
Major horse paper coming soon
Horse domestication is an important and controversial topic, in large part because it's intimately tied to the debate over the location of the Proto-Indo-European (PIE) homeland. Based on the currently available genetic and archaeological data, it seems likely that all modern domesticated horse breeds ultimately derive from the Pontic-Caspian steppe in Eastern Europe (see here and here).
In the interview linked to below (click on the image) horse expert Alan Outram reveals that a new paper will be published within months that will test this theory, and either confirm or debunk it.
 Outram also talks about the colonization of Central Asia during the Middle Bronze Age by groups from the west associated with the Sintashta culture. He says that this was probably an aggressive affair, akin to the more recent European colonization of North America, that may have pushed the Botai people, who were the indigenous inhabitants of the Kazakh steppe, and their horses far to the east. This, he suggests, might explain why the Przewalski horse of Mongolia appears to be derived from the Botai horse.
See also...
The mystery of the Sintashta people
Focus on Hittite Anatolia
Friendly Yeniseian steppe pastoralists
Outram also talks about the colonization of Central Asia during the Middle Bronze Age by groups from the west associated with the Sintashta culture. He says that this was probably an aggressive affair, akin to the more recent European colonization of North America, that may have pushed the Botai people, who were the indigenous inhabitants of the Kazakh steppe, and their horses far to the east. This, he suggests, might explain why the Przewalski horse of Mongolia appears to be derived from the Botai horse.
See also...
The mystery of the Sintashta people
Focus on Hittite Anatolia
Friendly Yeniseian steppe pastoralists
Friday, August 31, 2018
Focus on Hittite Anatolia
I computed a series of D-statistics on most of the currently available ancient samples from Central Anatolia - dating from almost the Epipaleolithic (Boncuklu_N) to the Hittite era (Anatolia_MLBA) - to try and get a better idea of who the Indo-European-speaking Hittites may have been. The full output as well as details about the key ancient samples used in this analysis are available here. See anything interesting? The most noteworthy statistics, I suppose, are those listed below, because they're significant (Z≥3) and organized chronologically.
 However, the thing to keep in mind in regards to D-statistics, and the very similar f4-statistics, when looking for signals of mixture is that they may or may not produce significant Z scores because of several reasons, such as the choice of the outgroup, the choice of the reference samples and the phylogenetic relationship between them, or even the type, quality and density of the data being used.
Perhaps ironically, the D-statistics above suggest that the Neolithic Central Anatolians (Boncuklu_N and Tepecik_Ciftlik_N) were more European-like than those from the Bronze Age, and I suspect that this is one of the main reasons why the idea of Eastern European admixture (from the Pontic-Caspian steppe and/or Balkans) in Hittites is currently being rejected by the geneticists working on the problem. But this dilemma is easy to explain away by the fact that the Neolithic samples carry much higher ratios of Anatolian Epipaleolithic hunter-gatherer admixture and also other types of ancestry shared with and/or closely related to European hunter-gatherers and early farmers.
In other words, I'd say that most of the statistics are being confounded by deep phylogenetic relationships, and thus aren't very useful for solving the Hittite problem. Interestingly, though, that relationship to Europe is reversed somewhat in the D-statistics involving Anatolia_EBA and Anatolia_MLBA, with the latter showing significantly higher affinity to Eastern European Hunter-Gatherers (EHG) and Minoans.
Thus, in my opinion, to get a more complete picture it's also useful to look for patterns in the statistics, even those that, strictly speaking, don't reach significance. One way to do that is with linear models. So here are a few linear models based on some of my D-statistics. The relevant datasheet is available here.
However, the thing to keep in mind in regards to D-statistics, and the very similar f4-statistics, when looking for signals of mixture is that they may or may not produce significant Z scores because of several reasons, such as the choice of the outgroup, the choice of the reference samples and the phylogenetic relationship between them, or even the type, quality and density of the data being used.
Perhaps ironically, the D-statistics above suggest that the Neolithic Central Anatolians (Boncuklu_N and Tepecik_Ciftlik_N) were more European-like than those from the Bronze Age, and I suspect that this is one of the main reasons why the idea of Eastern European admixture (from the Pontic-Caspian steppe and/or Balkans) in Hittites is currently being rejected by the geneticists working on the problem. But this dilemma is easy to explain away by the fact that the Neolithic samples carry much higher ratios of Anatolian Epipaleolithic hunter-gatherer admixture and also other types of ancestry shared with and/or closely related to European hunter-gatherers and early farmers.
In other words, I'd say that most of the statistics are being confounded by deep phylogenetic relationships, and thus aren't very useful for solving the Hittite problem. Interestingly, though, that relationship to Europe is reversed somewhat in the D-statistics involving Anatolia_EBA and Anatolia_MLBA, with the latter showing significantly higher affinity to Eastern European Hunter-Gatherers (EHG) and Minoans.
Thus, in my opinion, to get a more complete picture it's also useful to look for patterns in the statistics, even those that, strictly speaking, don't reach significance. One way to do that is with linear models. So here are a few linear models based on some of my D-statistics. The relevant datasheet is available here.


 Arguably, the most striking thing about these models is the position at the top of the graphs of the ancient populations from Central Asia and what is now Iran, and the gradually lower position of populations with progressively less of this type of ancestry. The most plausible explanation for this phenomenon is post-Boncuklu_N gene flow into Central Anatolia from the east, possibly as a continuation of something that was happening already since the Epipaleolithic, but becoming more intense during the Neolithic revolution, probably as a result of rapid population growth in and around the Fertile Crescent.
Indeed, I strongly suspect that one of the main reasons why we've been hearing so much lately about Iran as a likely candidate for the Indo-European homeland is this strong eastern signal in Bronze Age Anatolian DNA. If so, then this is likely to be a misunderstanding, because there are better explanations for it than the Indo-Europeans, such as the Hattians and Hurrians.
Another rather obvious outcome in my graphs is the relatively stronger affinity between the Bronze Age Anatolians and the ancient populations from Eastern Europe, including, and especially, those from the Pontic-Caspian steppe, compared to Tepecik_Ciftlik_N. In fact, looking at the Anatolia_EBA vs Tepecik_Ciftlik_N graph, I'd say that steppe admixture was already seeping into Central Anatolia during the Early Bronze Age.
If so, this is an important point that should be taken into account when modeling the ancestry of the Hittite era Anatolians. That's because if Anatolia_EBA already harbored some steppe ancestry, then we'd be shooting ourselves in the proverbial foot if we were to use it as the supposedly unadmixed reference population to try and determine whether Anatolia_MLBA was partly of steppe origin. Hence, to model the ancestry of Anatolia_MLBA, at least in the context of possible migrations from the steppe to Anatolia during the Bronze Age, it might be more useful to use Tepecik_Ciftlik_N as the likely unadmixed reference population.
Let's try that with qpAdm, first on the whole Anatolia_MLBA set, and then on one individual labeled MA2203, who, as far as I can tell, shows an elevated level of steppe ancestry in several different types of analyses. I chose Yamnaya_Kalmykia as the potential mixture source from the steppe because it's likely to be the closest available population in my dataset to the Eneolithic groups of the southern region of the Pontic-Caspian steppe.
Arguably, the most striking thing about these models is the position at the top of the graphs of the ancient populations from Central Asia and what is now Iran, and the gradually lower position of populations with progressively less of this type of ancestry. The most plausible explanation for this phenomenon is post-Boncuklu_N gene flow into Central Anatolia from the east, possibly as a continuation of something that was happening already since the Epipaleolithic, but becoming more intense during the Neolithic revolution, probably as a result of rapid population growth in and around the Fertile Crescent.
Indeed, I strongly suspect that one of the main reasons why we've been hearing so much lately about Iran as a likely candidate for the Indo-European homeland is this strong eastern signal in Bronze Age Anatolian DNA. If so, then this is likely to be a misunderstanding, because there are better explanations for it than the Indo-Europeans, such as the Hattians and Hurrians.
Another rather obvious outcome in my graphs is the relatively stronger affinity between the Bronze Age Anatolians and the ancient populations from Eastern Europe, including, and especially, those from the Pontic-Caspian steppe, compared to Tepecik_Ciftlik_N. In fact, looking at the Anatolia_EBA vs Tepecik_Ciftlik_N graph, I'd say that steppe admixture was already seeping into Central Anatolia during the Early Bronze Age.
If so, this is an important point that should be taken into account when modeling the ancestry of the Hittite era Anatolians. That's because if Anatolia_EBA already harbored some steppe ancestry, then we'd be shooting ourselves in the proverbial foot if we were to use it as the supposedly unadmixed reference population to try and determine whether Anatolia_MLBA was partly of steppe origin. Hence, to model the ancestry of Anatolia_MLBA, at least in the context of possible migrations from the steppe to Anatolia during the Bronze Age, it might be more useful to use Tepecik_Ciftlik_N as the likely unadmixed reference population.
Let's try that with qpAdm, first on the whole Anatolia_MLBA set, and then on one individual labeled MA2203, who, as far as I can tell, shows an elevated level of steppe ancestry in several different types of analyses. I chose Yamnaya_Kalmykia as the potential mixture source from the steppe because it's likely to be the closest available population in my dataset to the Eneolithic groups of the southern region of the Pontic-Caspian steppe.




Anatolia_MLBA Seh_Gabi_ChL 0.200±0.043 Tepecik_Ciftlik_N 0.659±0.033 Yamnaya_Kalmykia 0.141±0.022 chisq: 11.425 tail prob: 0.408405 Full output MA2203 Seh_Gabi_ChL 0.179±0.065 Tepecik_Ciftlik_N 0.622±0.049 Yamnaya_Kalmykia 0.199±0.036 chisq: 12.914 tail prob: 0.299004 Full outputPlease note, however, that these mixture models are based on f4-statisctics. So, obviously, they're going to be affected by the same factors as described above that affect f4-statistics. Hence, despite the seemingly statistically sound output, the steppe admixture that you see there might not actually be admixture from the steppe. In fact, there's a good reason why I'm not shouting from the rooftops that I've just uncovered the presence of steppe ancestry in Bronze Age Anatolia, and thus confirmed the steppe or kurgan hypothesis positing that the Hittite and indeed Indo-European homeland was located in the Pontic-Caspian steppe. That's because I used a mixed bag of UDG-treated capture data and non-UDG-treated shotgun data. This is known to be a serious problem, which can skew the results of even the most robust analyses, and produce spurious statistics and Z scores. Nevertheless, I'm reasonably confident that my findings will eventually be confirmed with more and higher quality data from ancient Anatolia. Let's wait and see. See also... Late PIE ground zero now obvious; location of PIE homeland still uncertain, but...
Friday, August 24, 2018
Global25 workshop 3: genes vs geography in Northern Europe
To produce the intra-North European Principal Components Analysis (PCA) plot below, download this datasheet, plug it into the PAST program, which is freely available here, then select all of the columns by clicking on the empty tab above the labels, and choose Multivariate > Ordination > Principal Components or Discriminant Analysis. This is what you should end up with...

 I'd say that the result more or less resembles a geographic map of Northern Europe. Of course, if you're in the possession of your own personal Global25 coordinates, you can add yourself to this plot to check whether your position matches your geographic origin.
Please keep in mind, however, that the vast majority (>90%) of your ancestry must be from north of the Alps, Balkans and Pyrenees to obtain a sensible outcome. Also please ensure that all of the columns in the datasheet are filled out correctly, including the group column, otherwise your position on the plot will be skewed.
See also...
Global25 workshop 1: that classic West Eurasian plot
Global25 workshop 2: intra-European variation
Global25 workshop 4: a neighbour joining tree
Getting the most out of the Global25
Genetic ancestry online store (to be updated regularly)
I'd say that the result more or less resembles a geographic map of Northern Europe. Of course, if you're in the possession of your own personal Global25 coordinates, you can add yourself to this plot to check whether your position matches your geographic origin.
Please keep in mind, however, that the vast majority (>90%) of your ancestry must be from north of the Alps, Balkans and Pyrenees to obtain a sensible outcome. Also please ensure that all of the columns in the datasheet are filled out correctly, including the group column, otherwise your position on the plot will be skewed.
See also...
Global25 workshop 1: that classic West Eurasian plot
Global25 workshop 2: intra-European variation
Global25 workshop 4: a neighbour joining tree
Getting the most out of the Global25
Genetic ancestry online store (to be updated regularly)


Wednesday, August 8, 2018
The South Asian cline that no longer exists
Before the Indo-Europeans and Austroasiatics got to South Asia, probably well within the last 4,000 years, it's likely that all of the genetic variation in the region basically sat along a genetic cline devoid of any Bronze Age steppe and Southeast Asian ancestry, like the one in the Principal Component Analysis (PCA) below running from the Paniya to the "Indus Periphery" ancient sample Shahr_I_Sokhta BA2.
 Note that almost all of the South Asian populations, including the Iron Age (IA) Swat Valley groups, are clearly peeling away from the said cline towards the Tajiks, in other words towards Central Asia. This is a reflection of the widespread presence of Sintashta-related steppe admixture among South Asians, especially those speaking Indo-European languages. Moreover, the Bangalis and Burushos are being pushed towards the top left of the plot as a result of East Asian-related ancestry. In the case of the former, this is largely due to gene flow from Austroasiatic groups.
It'll be interesting to see how ancient Harappans behave in this analysis. I'm betting that they'll be very similar to the Indus Periphery trio, although judging by the latest press report on the topic (see here), the Harappan samples from Rakhigarhi might be shifted much closer to the Paniya as a result of a higher ratio of indigenous South Asian ancestry.
The PCA is based on my Global25 test. If you're South Asian and in the possession of Global25 coordinates, you can add yourself to this plot using the datasheet available here. Plug the datasheet into the PAST program (freely available here), select all of the columns, and go Multivariate > Ordination > Principal Components (PCA).
Update 10/08/2018: I managed to almost reproduce my PCA with a graph based on D-stats of the form D(Mbuti,X)(Onge,Ganj_Dareh_N)/D(Mbuti,X)(Ganj_Dareh_N,Sintashta_MLBA). Admittedly, Gonur2_BA didn't want to cooperate by pushing slightly up and away from the ghost South Asian cline. But this may have been due to a lack of data or perhaps minor admixture (keep in mind that this sample is actually from Turkmenistan and not South Asia). However, combining all three of the Indus_Periphery individuals worked well enough. The relevant datasheet is available here.
Note that almost all of the South Asian populations, including the Iron Age (IA) Swat Valley groups, are clearly peeling away from the said cline towards the Tajiks, in other words towards Central Asia. This is a reflection of the widespread presence of Sintashta-related steppe admixture among South Asians, especially those speaking Indo-European languages. Moreover, the Bangalis and Burushos are being pushed towards the top left of the plot as a result of East Asian-related ancestry. In the case of the former, this is largely due to gene flow from Austroasiatic groups.
It'll be interesting to see how ancient Harappans behave in this analysis. I'm betting that they'll be very similar to the Indus Periphery trio, although judging by the latest press report on the topic (see here), the Harappan samples from Rakhigarhi might be shifted much closer to the Paniya as a result of a higher ratio of indigenous South Asian ancestry.
The PCA is based on my Global25 test. If you're South Asian and in the possession of Global25 coordinates, you can add yourself to this plot using the datasheet available here. Plug the datasheet into the PAST program (freely available here), select all of the columns, and go Multivariate > Ordination > Principal Components (PCA).
Update 10/08/2018: I managed to almost reproduce my PCA with a graph based on D-stats of the form D(Mbuti,X)(Onge,Ganj_Dareh_N)/D(Mbuti,X)(Ganj_Dareh_N,Sintashta_MLBA). Admittedly, Gonur2_BA didn't want to cooperate by pushing slightly up and away from the ghost South Asian cline. But this may have been due to a lack of data or perhaps minor admixture (keep in mind that this sample is actually from Turkmenistan and not South Asia). However, combining all three of the Indus_Periphery individuals worked well enough. The relevant datasheet is available here.

 See also...
Global25 workshop 1: that classic West Eurasian plot
Global25 workshop 2: intra-European variation
Global25 workshop 3: genes vs geography in Northern Europe
Global25 workshop 4: a neighbour joining tree
Getting the most out of the Global25
Genetic ancestry online store (to be updated regularly)
See also...
Global25 workshop 1: that classic West Eurasian plot
Global25 workshop 2: intra-European variation
Global25 workshop 3: genes vs geography in Northern Europe
Global25 workshop 4: a neighbour joining tree
Getting the most out of the Global25
Genetic ancestry online store (to be updated regularly)



Subscribe to:
Posts (Atom)